Penjelasan Lengkap Genetika Bakteri – Ilmu genetika mendefinisikan dan menganalisis keturunan dan perubahan pengaturan dari berbagai fungsi fisiologis yang membentuk karakter organisme. Unit keturunan disebut gen yang merupakan suatu segmen DNA yang nukleotidanya membawa informasi karakter biokimia atau fisiologis tertentu. Pendekatan tradisional pada genetika telah mengidentifikasikan gen sebagai dasar kontribusi karakter fenotip atau karakter dari keseluruhan stuktural dan fisiologis dari suatu sel atau organisme, karakter fenotip seperti warna mata pada manusia atau resistensi terhadap antibiotik pada bakteri, pada umumnya di amati pada tingkat organisme. Dasar kimia untuk variasi dalam fenotip atau perubahan urutan DNA dalam suatu gen atau dalam organisasi gen.
Penelaahan tentang genetika pertama kali dilakukan oleh seorang ahli botani bangsa Austria, Gregor Mendel pada tanaman kacang polongnya. Pada tahun 1860-an ia menyilangkan galur-galur kacang polong. Hasilnya antara lain terjadi perubahan-perubahan pada warna,bentuk, ukuran, dan sifat-sifat lain dari kacang polong tersebut. Penelitian inilah ia mengembangkan hukum-hukum dasar kebakaan. Hukum kebakaan berlaku umum bagi semua bentuk kehidupan. Hukum-hukum mendel berlaku manusia dan juga organisme percobaan dahulu amat populer dalam genetika, yakni lalat buah Drosophila. Namun sekarang, percobaan-percobaan ilmu kebakaan dengan menggunakan bakteri Escherichia coli. Bakteri ini dipilih karena paling mudah dipelajari pada taraf molekuler sehingga merupakan organisme pilihan bagi banyak ahli genetika. Hal ini membantu perkembangan bidang genetika mikroba. Jasad renik yang di pelajari dalam bidang genetika mikroba meliputi bakteri, khamir, kapang, dan virus.
Genetika mikroba tradisional terutama berdasarkan pada pengamatan atau observasi perkembangan secara luas. Variasi fenotip telah diamati berdasar kemampuan gen untuk tumbuh dibawah kondisi terseleksi, misalnya bakteri yang mengandung satu gen yang resisten terhadap ampisilin dapat dibedakan dari bakteri kekurangan gen selama pertumbuhannya dalam lingkungan yang mengandung antibiotik sebagai suatu bahan penyeleksi. Catatan bahwa seleksi gen memerlukan ekspresinya dibawah kondisi yang tepat dapat diamati pada tingkat fenotif. Genetika bakteri mendasari perkembangan rekayasa genetika, suatu teknologi yang bertanggung jawab terhadap perkembangan di bidang kedokteran.
APA ITU GENOM BAKTERI
Unit herediter pada bakteri disebut genom bakteri (gen bakteri). Sebuah sel memiliki DNA yang merupakan materi genetik dan bersifat herediter pada seluruh sistem kehidupan. Genom adalah set lengkap materi genetik (DNA) yang dimiliki suatu organisme dan terorganisasi menjadi kromosom. Gen bakteri terdapat dalam molekul DNA tunggal (haploid). Kromosom bakteri yang terdiri dari DNA berbentuk sirkuler, panjangnya ± 1mm, beratnya 2-3% dari berat kering satu sel, disusun sekitar 4 juta kpb DNA, makromolekul yang sangat banyak ini dikemas agar tidak berubah dalam bentuk superkoil (± 70-130 superkoil domain). DNA memiliki berat molekul yang tinggi karena terdiri dari heteropolimer dari deokdiribonukleotida purin (adenin dan guanin) dan pirimidin (sitosin dan timin). Selain itu dikenal pula adanya materi genetik di luar kromosom (ekstra kromosomal) yang disebut plasmid.
Plasmid tersebar luas dalam populasi bakteri. Plasmid terdiri dari beberapa – 100 kpb, beratnya ± 1-3 % dari kromosom –bakteri, berada bebas dalam sitoplasma bakteri. Kadang-kadang dapat bersatu dengan kromosom bakteri. Dapat berpindah dan dipindahkan dari satu spesies ke spesies lain. Jumlahnya dapat mencapai 30 atau dapat bertambah karena mutasi. Meskipun bakteri bersifat haploid, transmisi gen dari satu generasi ke generasi berikutnya berlangsung secara linier, sehingga pada setiap siklus pembelahan sel, sel anaknya menerima satu set gen yang identik dengan sel induknya.
Ada dua fenomena biologi pada konsep hereditas:
- Hereditas yang bersifat stabil dimana generasi berikutnya yang terbentuk dari pembelahan satu sel mempunyai sifat yang identik dengan induknya.
- Variasi genetik yang mengakibatkan adanya perbedaan sifat generasi berikut dan sel induknya akibat peristiwa genetik tertentu, misalnya mutasi.
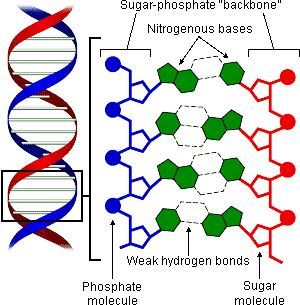
Watson dan Crick, dengan sinar X menemukan bahwa struktur DNA terdiri dari dua rantai poliribonukleotida yang dihubungkan satu sama lain oleh ikatan hidrogen antara purin di satu rantai dengan pirimidin di rantai lain, dalam keadaan antiparalel, dan disebut sebagai struktur double helix. Ikatan hidrogen ini hanya dapat menghubungkan Adenin (6 aminopurin) dengan Timin (2,4 dioksi 5 metil pirimidin) dan antara Guanin (2 amino 6 oksipurin) dengan Sitosin (2 oksi 4 amino pirimidin). Singkatnya pasangan basa pada suatu sekuens DNA adalah A-T dan S-G. Karena adanya sistem berpasangan demikian, maka setiap rantai DNA dapat dijadikan cetakan/template untuk membangun rantai DNA yang komplementer. Waktu terjadinya proses replikasi DNA dalam pembelahan sel, molekul DNA dari sel anaknya terdiri dari satu rantai DNA yang komplememter tapi dibuat baru, dengan kata lain, pemindahan materi genetik dari satu generasi ke generasi berikutnya adalah dengan cara semikonservatif.
 |
Fungsi primer DNA pada hakikatnya adalah sebagai sumber perbekalan informasi genetik yang dimiliki oleh sel induk. Proses replikasi di kerjakan dengan amat lengkap sehingga sel anaknya mendapatkan pula informasi genetik yang lengkap, sehingga terjadi kesetabilan genetik dalam suatu populasi mikroorganisme. Satu benang kromosom biasanya terdiri dari lima juta pasangan basa dan terbagi atas segmen atau sekuens asam amino tertentu yang akan membentuk stuktur protein. Protein ini kemudian menjadi enzim-enzim, komponen membran sel dan struktur sel yang lain yang secara keseluruhan menentukan karakter dari sel itu.
DNA BAKTERI
REPLIKASI

- Replikasi DNA dimulai pada tempat-tempat khusus yang disebut pangkal replikasi (origin of replication, ori)
- Kromosom prokariot hanya mempunyai satu ori, sedangkan kromosom eukariot mempunyai beberapa ori
- Setiap untai DNA baru disintesis melalui dua mekanisme:
- Sintesis kontinyu oleh DNA polimerase III dengan arah 5’ ke 3’ menghasilkan Leading strand
- Sintesis diskontinyu oleh DNA polimerase III dengan arah 5’ ke 3’ menghasilkan fragmen pendek (± 1000 nukleotida) yang disebut fragmen Okazaki pada Lagging strand
TRANSKRIPSI
- RNA polimerase mengikat urutan DNA yang disebut promotor.
- RNA polymerase mensintesis RNA diarahkan oleh satu untai DNA cetakan
- DNA cetakan yang telah ditranskripsi dipilin kembali
- Sintesis RNA berakhir ketika RNA polimerase mencapai urutan DNA yang disebut terminator.
- Molekul RNA baru dan RNA polimerase terlepas dari DNA cetakan

TRANSLASI

2. Elongasi
Proses elongasi terjadi saat tRNA dengan antikodon dan asam amino yang tepat masuk ke tempat A. Akibatnya kedua tempat di ribosom terisi, lalu terjadi ikatan peptide antara kedua asam amino. Ikatan tRNA dengan Nformil metionin lalu lepas, sehingga kedua asam amino yang berangkai berada pada tempat A. Ribosom kemudian bergeser sehingga asam amino-asam amino-tRNA berada pada tempat P dan tempat A menjadi kosong. Selanjutnya tRNA dengan antikodon yang tepat dengan kodon ketiga akan masuk ke tempat A, dan proses berlanjut seperti sebelumnya..
3. Terminasi.
Proses translasi akan berhenti bila tempat A bertemu kodon akhir yaitu UAA, UAG, UGA. Kodon-kodon ini tidak memiliki tRNA yang membawa antikodon yang sesuai. Selanjutnya masuklah release factor (RF) ke tempat A dan melepaska rantai polipeptida yang terbentuk dari tRNA yang terakhir. Kemudian ribosom berubah menjadi sub unit kecil dan besar.
Baca juga :
- Bakteri penghasil antibiotik
- Bakteri gram positif dan negatif, apa bedanya?
- Peran bakteri pada industri pangan
MUTASI
Mutasi mengarah pada suatu perubahan senyawa kimia pada DNA. Mutan merupakan individu yang mengalami perubahan pada satu atau lebih basa DNAnya: perubahan ini dapat diwariskan dan irreversibel (kecuali terjadi mutasi-balik ke urutan awal).
Kerusakan gen tersebut dapat disebabkan oleh:
- Perubahan pada proses transkripsi
- Perubahan pada urutan asam amino dari protein yang merupakan produk gen.
Mutasi adalah perubahan di dalam rangkaian nukleotida suatu gen. Mutasi menimbulkan ciri genetik yang baru atau genotif berubah. Sel atau organisme yang menimbulkan efek mutasi disebut mutan. Mutasi pada gen akan menyebabkan produk protein yang dihasilkan.
Mutasi melibatkan sejumlah gen pada DNA bakteri: beberapa mutasi tidak pernah dideteksi karena tergantung pada mutasi mempengaruhi suatu fungsi yang dapat dikenali (contoh, penyebab resistensi antibiotik). Dan yang lain dapat mematikan sehingga tidak terdeteksi. Mutasi dikelompokkan berdasarkan :
1. Ukuran
Mutasi titik. suatu perubahan pada sebagian kecil segmen DNA; biasanya yang terjadi pada suatu nukleotida tunggal atau pasangan nukleotida.
- Samesense (silent) mutasi: perubahan pada suatu kodon (biasanya pada posisi ke tiga) yang gagal untuk memindahkan asam amino spesifik dari tempat yang tidak mengalami mutasi.
- Nonsense mutasi : suatu pemendekan produk protein , pada signal rantai-terminasi
- Missense mutasi: suatu perubahan urutan asam amino dengan asam amino yang mengalami salah cetak ditempatkan pada rantai polipeptida
- Frameshift mutasi: suatu pergeseran reading frame, menghasilkan sejumlah kodon missense dan nonsense melalui sisa cistron
- “Gross Mutasi”: perpindahan yang melibatkan lebih dari satu pasangan nukleotida, dapat memasukkan gen, kromosom, atau rangkaian kromosom (pada eukariota).
2. Kualitas
Struktural mutasi merupakan perubahan pada nukleotida yang mengandung gen :
- Substitusi mutasi: penggantian satu nukleotida untuk yang lainnya, dapat dibedakan menjadi, Transisi: pertukaran dalam satu purindengan satu pirimidin atau sebaliknya (contoh, GC menjadi AT); Transversi: perubahan pada purin/pirimidin (contoh, GC menjadi CG).
- Delesi mutan: kehilangan beberapa bagian suatu gen.
- Insersi mutan: penambahan satu atau lebih ekstra nukleotida terhadap suatu gen.
- ”Rearragement Mutasi” merupakan perubahan lokasi suatu gen dalam genom, sering diikuti oleh efek posisi.
- Dalam suatu gen: dua mutasi dalam gen yang sama fungsinya dapat menghasilkan efek yang berbeda, tergantung pada terjadinya apakah pada posisi cis atau trans.
- Sejumlah gen tiap kromosom: efek fenotip berbeda dapat dihasilkan jika sejumlah gen yang mengalami replikasi nonequivalen pada kromosom homolog (eukariota).
- Pergerakan lokus gen dapat menghasilkan fenotip baru, khususnya ketika gen dipindahkan dekat heterokromatin (eukariot).
3. Asal
Mutasi spontan awalnya tidak diketahui, sering disebut background mutation. Kontrol genetik mutabilitas beberapa gen yang diketahui dapat disebabkan oleh “mutator gen” lain. Mutasi spontan dapat dibedakan menjadi 1) mutasi spesifik yang pengaruhnya terbatas pada satu lokus dan 2) mutasi nonspesifik secara simultan mempengaruhi pada beberapa lokus. Mutasi terinduksi dipengaruhi oleh keadaan lingkungan yang tidak normal, misalnya: Radiasi pengion (perubahan valensi senyawa kimia melalui penembahan elektron yang dihasilkan oleh proton, neutron, atau oleh sinar X. Radiasi nonpengion penambahan tingkat energi atom (eksitasi), yang membuatnya kurang stabil (contoh, radiasi UV, panas), radiasi UV sering menghasilkan dimer timin, contoh, ikatan timin di antara dua rantai yang sama. Mutagen senyawa kimia (senyawa kimia yang meningkatkan mutabilitas gen) dapat dibedakan menjadi: Salah cetak mutan meningkat selama replikasi DNA (contoh, mutagen analog basa dengan sifat kimia yang sama dengan basa asam nukleat dapat masuk karena kesalahan, akridin penyebab penambahan mutasi tunggal atau delesi kemungkinan karena interkalasi di antara dua urutan basa). Perubahan gen langsung dihasilkan pada DNA nonreplicating, contohnya asam nitrat oleh deaminasi secara langsung merubah adenin menjadi hipoksantin dan sitosin menjadi urasil.
Mutagenesis
Mutagenesis merupakan suatu teknik biologi molekuler di mana suatu mutasi diciptakan pada suatu bagian molekul DNA tertentu, yang dikenal sebagai plasmid. Mutagenesis adalah proses biologis yang sangat mendasar. Secara tradisional, percobaan pada mutagenesis bakteri telah dilakukan menggunakan kultur pertumbuhan bakteri karena keyakinan bahwa hanya tumbuh sel-sel (mengandung DNA yang bereplikasi) yang bisa berubah. Namun, akhir-akhir ini menunjukkan bahwa bakteri rentan terhadap mutagenesis tidak hanya selama pertumbuhan, tetapi juga ketika mengalami tekanan seleksi tersebut yang mencegah pertumbuhan, tetapi tidak mematikan.
Dalam mutagenesis yang tidak mematikan seleksi secara bebas disebut `adaptif’ sejak mutasi yang biasanya ditemukan memungkinkan sel untuk melawan tekanan dan melanjutkan pertumbuhan. Namun, banyak pekerja lebih suka menggunakan istilah fase stasioner mutagenesis sebagai gantinya. Proses ini telah banyak diperdebatkan sejak 1988 dan banyak ulasan pada subjek yang tersedia.
Mekanisme dasar:
- Mensintesis DNA yang di dalamnya terdapat bagian yang ingin dimutasi.
- Hasil sintesis ini harus dihibridisasi dengan DNA lain dari gen yang diinginkan.
- Fragmen tersebut diperluas lagi oleh DNA polimerase.
- Molekul yang diperoleh akan diadaptasikan ke dalam sel inang dan dikloning.
- Pemilihan mutan.

Mutagen
Bahan-bahan yang menyebabkan terjadinya mutasi disebut mutagen. Mutagen terbagi menjadi tiga, yaitu:
1. Mutagen bahan kimia
Mutagen bahan kimia, contohnya adalah kolkisin dan zat digitonin. Kolkisin adalah zat yang dapat menghalangi terbentuknya benang-benang spindel pada proses anafase dan dapat menghambat pembelahan sel pada anafase. Mutagen bahan kimia dapat menimbulkan mutasi melalui beberapa cara. Gugusan alkil aktif dari bahan mutagen kimia dapat ditransfer ke molekul lain pada posisi dimana kepadatan elektron cukup tinggi seperti PHOSPHATE GROUPS dan juga molekul PURINE dan PYRIMIDINE yang merupakan penyusun struktur deoxyribonucleic acid (DNA). Seperti diketahui umum, DNA merupakan struktur kimia yang membawa gen. Basa-basa yang menyusun struktur DNA terdiri dari ADENINE, GUANINE, THYIMINE, dan CYTOSINE. Adenine dan guanine merupakan basa bercincin ganda (DOUBLE-RING BASES) disebut PURINES, sedangkan thymine dan cytosine bercincin tunggal (SINGLE-RING BASES) disebut PYRIMIDINES. Struktur molekul DNA berbentuk pilitan ganda (DOUBLE HELIX) dan tersusun atas pasangan spesifik ADENINE-THYMINE dan GUANINE-CYTOSINE. Contoh mutasi yang paling sering ditimbulkan oleh mutagen kimia adalah perubahan basa pada struktur DNA yang mengarah pada pembentukan 7-ALKYL GUANINE.
2. Mutagen bahan fisika
Mutagen bahan fisika, contohnya sinar ultraviolet, sinar radioaktif, dan lain-lain. Sinar ultraviolet dapat menyebabkan kanker kulit. Mutagen fisika bersifat sebagai radiasi pengion (IONIZING RADIATION) yang dapat melepas energi (ionisasi), begitu melewati atau menembus materi. Mutagen fisika termasuk diantaranya sinar-X, radiasi gamma, radiasi beta, neutron, dan partikel dari aselerators sudah umum digunakan dalam pemuliaan tanaman. Karakteristik untuk masing-masing jenis radiasi disajikan dalam tabel di bawah ini. Begitu materi reproduksi tanaman diradiasi, proses ionisasi akan terjadi dalam jaringan dan dapat menyebabkan perubahan pada jaringan itu sendiri, sel, genom, kromosom, dan DNA atau gen. Perubahan yang ditimbulkan pada tingkat genom, kromosom, dan DNA atau gen dikenal dengan istilah mutasi (MUTATION).
3. Mutagen bahan biologi
Diduga virus dan bakeri dapat menyebabkan terjadinya mutasi. Bagian virus yang dapat menyebabkan terjadinya mutasi adalah DNA-nya.
Analisis genetik bakteri memerlukan adanya mutan-mutan, yang secara garis besar dapat dikelompokkan ke dalam tiga kategori utama:
- Mutan resisten antibiotik, yaitu mutan yang mampu tumbuh pada medium yang mengandung antibiotik tertentu seperti streptomisin (Str), ampisilin (Amp), dan tetrasiklin (Tet). Sel-sel bakteri alami (wild type) tidak mampu tumbuh pada medium yang mengandung salah satu antibiotik tersebut. Sebaliknya, mutan resisten ampisilin (Amp-r) misalnya, dapat tumbuh pada medium dengan antibiotik ampisilin.
- Mutan nutrisional, yaitu mutan yang tidak dapat menyintesis nutrien-nutrien esensial seperti yang dilakukan oleh sel-sel bakteri alami sehingga mutan ini tidak mampu tumbuh kecuali dengan pemberian nutrien yang diperlukan ke dalam medium pertumbuhannya. Mutan bakteri semacam ini dikatakan sebagai auksotrof untuk nutrien tertentu. Sebagai contoh, auksotrof metionin tidak dapat tumbuh pada medium minimal, yang hanya mengandung garam-garam anorganik serta sumber atom karbon atau energi. Mutan ini baru akan tumbuh apabila ke dalam medium tersebut ditambahkan metionin.
- Mutan sumber karbon, yaitu mutan yang tidak dapat menggunakan zat-zat tertentu sebagai sumber karbon atau energi. Sebagai contoh, mutan Lac- tidak dapat menggunakan gula laktosa untuk pertumbuhannya sehingga tidak akan membentuk koloni pada medium minimal yang mengandung laktosa sebagai satu-satunya sumber karbon.
TRANSFER GENETIK BAKTERI
Perpindahan gen merupakan suatu kegiatan yang dilakukan bakteri dengan mengirimkan informasi genetik (DNA) dari sel donor ke sel resipien. Kegiatan perpindahan gen ini ada tiga yakni :
a. Transformasi
Transformasi pertama kali ditemukan oleh Frederick Griffith pada tahun 1928. Dia mempelajari transformasi satu tipe Streptococcus pneumoniae menjadi tipe yang berbeda. S. pneumoniae dibagi menjadi 100 tipe lain yang berbeda atas dasar perbedaan kimia pada kapsulnya. Jadi, tipe 1 menghasilkan kapsul yang berbeda dengan tipe 2, dan seterusnya.
Transformasi ialah proses pemindahan DNA bebas sel yang mengandung sejumlah informasi genetik (DNA) dari satu sel ke sel lainnya. DNA tersebut diperoleh dari sel donor melalui lisis sel alamiah atau dengan cara ekstraksi kimiawi. Begitu fragmen DNA dari sel donor tertangkap oleh sel resipien, maka terjadilah rekombinasi.

Manfaat yang didapat dari transformasi gen pada bakteri adalah :

c. Transduksi

PERBAIKAN KERUSAKAN DNA
Sebagian DNA yang rusak diperbaiki sebelum menyebabkan perubahan menjadi mutasi genetik yang diturunkan. Semua organisme, termasuk bakteri memiliki jalur yang benar-benar kompleks dan efisien untuk memperbaiki berbagai kerusakan pada DNA. Sedangkan E. coli tipe-liar secara efisien dapat bertahan dari dosis UV yang sangat tinggi, mutan tanpa kemampuan untuk memperbaiki kerusakan karena UV dibunuh oleh dimer pirimidin tunggal. Perbaikan dimer tersebut menggambarkan strategi umum dari jalur perbaikan DNA utama. Perlu dicatat bahwa pada sebagian besar jalur perbaikan kemungkinan hanya karena dobel-heliks alami DNA.
Cetakan rantai yang rusak tersebut biasanya digunakan untuk mengganti informasi yang hilang pada saat DNA rusak. Perbaikan DNA yang rusak dapat dilakukan dengan cara :
a. Pembalikan Langsung Dari Kerusakan (Direct Reversal of Damage)
Terdapat dua jalur yang sudah difahami untuk memperbaiki kerusakan DNA secara langsung dengan suatu mekanisme pembalikan tunggal. Salah satunya adalah fotoreaktivasi, melibatkan suatu flavoprotein yang disebut fotoliase yang merubah dimer siklobutan kembali menjadi unsur pokoknya yaitu pirimidin. Peran fotoliase membutuhkan cahaya, jadi jalur ini dapat dihentikan dengan penginkubasian bakteri dalam kegelapan. Fotoliase E. coli dikode oleh gen phr, dan aktivitasnya yang sama dapat dideteksi pada beberapa organisme lain.
Mekanisme pembalikan langsung kedua berperan pada “alkyl-substituted DNA” dan dikatalisis oleh suatu kelompok enzim yang disebut metiltransferase. Gen ada E. coli pengkode suatu protein yang dapat menerima grup metil secara langsung dari DNA, dengan cara demikian dapat membalikkan paling tidak tiga macam kerusakan alkilasi yang berbeda (termasuk 6-metilguanin, lesi premutagenik utama). Grup metil ditransfer ke satu dari dua residu sistein pada protein. Secara menyolok, residu sistein termetilasi tidak dapat dibalikkan, jadi metiltransferase berperan dalam suatu cara “bunuh diri”.
Protein ada juga mengendalikan respon adaptif, suatu jaringan pengatur yang dilibatkan pada perbaikan kerusakan alkilasi. Alkilasi satu dari residu sistein merubah protein ada menjadi suatu positif regulator yang mengaktifkan transkripsi paling sedikit tiga operon. Satu dari operon pengkode ada oleh karena itu alkilasi protein ada menghasilkan banyak protein ada. Operon kedua termasuk suatu “methylated-basespesific DNA glycosylase” yang menghancurkan kerusakan menjadi suatu jalur perbaikan eksisi. Minimal dua jalur dari perbaikan alkilasi diinduksi oleh respon adaptif, memulai suatu peningkatan yang besar dalam pertahanan hidup sesudah pemberiandengan zat penyebab alkilasi.
b. Perbaikan Eksisi/pengeluaran (Excision Repair)
Jalur perbaikan exsisi lazim dalam semua organisme yang diteliti dan merupakan mekanisme umum terpenting dari perbaikan DNA. Seluruh proses perbaikan eksisi melibatkan pemotongan satu rantai DNA dekat kerusakan, pengeluaran bagian rantai yang mengandung basa yang rusak, resintesis melalui gap, dan selanjutnya ligasi/menyambung untuk menyimpan heliks ganda yang utuh. Serangkaian kompleks reaksi tersebut membutuhkan bantuan beberapa protein yang sama yang dilibatkan pada replikasi genom, dengan DNA polimerase I biasanya menyelenggarakan sintesis pengganti pada E. coli. Fokus di sini ialah pada reaksi perbaikan eksisi awal, dan dikatalisis oleh beberapa kompleks protein yang mengenali bentuk kerusakan tertentu.
Proses perbaikan eksisi dari dimer siklobutan terinduksi-UV pada E. coli dimulai dengan peran suatu kompleks endonuklease yang dikode oleh tiga gen, uvrA, uvrB, dan uvrc. Nuklease tersebut mengenal lesi DNA, memotong/insisi rantai yang rusak pada setiap sisi lesi, dengan cara demikian mengeluarkan aligosakarida dari basa 12 atau 13 yang mengandung lesi. Resintesis menggunakan rantai komplemen sebagai cetakan dan penggabungan akhir melengkapi proses perbaikan tersebut. Bukti yang ada sekarang enzim tersebut dapat berfungsi bersama, dengan terkoordinir reaksi insisi dan resintesis. Nuklease uvrABC mengenali kerusakan yang terinduksi-UV dan secara kovalen terserang daerah lesi, kemungkinan dengan penggunaan suatu gross pertubation pada DNA.
c. Perbaikan Bypass dan Sistem SOS
Suatu rangkaian khusus dari reaksi perbaikan dibawa oleh protein yang secara khusus diinduksi dalam merespon bentuk kerusakan DNA tertentu. Dalam respon terhadap kerusakan DNA, E. coli menginduksi suatu regulon global yang mempengaruhi bermacam-macam proses termasuk perbaikan DNA, mutagenesis, rekombinasi, dan pembelahan sel. Dua kunci elemen pengatur pada regulon SOS ini adalah produk dari gen recA dan lexA. Protein recA merupakan enzim multifungsional yang terlibat dalam beberapa reaksi penting yang melibatkan DNA dan juga berfungsi sebagai protease pada saat terjadi kerusakan DNA. Proteolisis terinduksi melakukan degradasi protein lexA, yang secara normal merupakan repessor semua operon pada regulon SOS. Satu dari gen tersebut dikendalikan lexA merupakan recA dengan sendirinya, dan oleh karena itu sintesis protein recA diinduksi oleh kerusakan DNA dalam suatu putaran balik (feedback loop). Aktivasi fungsi protease dari protein recA tidak secara lengkap difahami. Bukti.
yang sekarang ada menunjukkan bahwa beberapa sinyal yang cukup, dan termasuk lesi DNA tertentu (seperti fotoproduk UV), oligosakarida DNA pendek yang dihasilkan dari perbaikan DNA, dan rantai-tunggal DNA yang dihasilkan dari penghambatan replikasi kromosom.Hal ini terhitung untuk fakta bahwa respon SOS diinduksi oleh berbagai pemberian mutagenik dan oleh kondisi nonmutagenik yang menghentikan replikasi.
Baca juga :
Semoga bermanfaat